Introduction
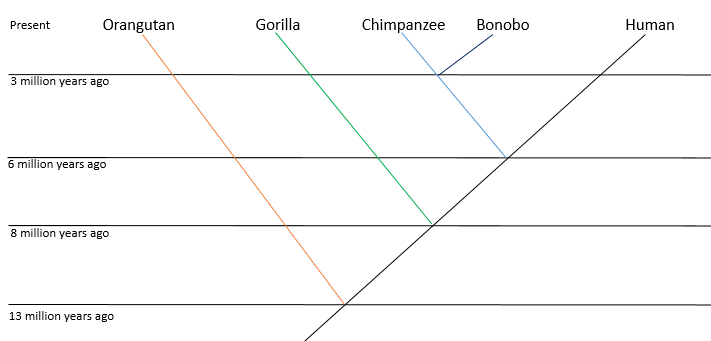
The behaviour of the closest living relatives to modern humans can give a tentative insight into how early humans may have behaved. Through observing primates’ cognitive abilities, social structure, cultural behaviour and expression of compassion and morality, one can begin to understand the behaviour of early humans and perhaps suggest behavioural parallels in human evolution to that of modern primates. However, there are limitations to using primate analogies; despite being the closest living relatives to modern humans, at least 6 million years of evolution separates both modern humans and modern primates from the last common ancestor (Figure 1), and both species diverged in vastly different paths. Thus, it cannot be assumed that modern primates have not evolved from the last common ancestor, and consequently this limits the extent to which modern primate behaviour can be applied to early modern human behaviour.
Cognition
Advanced cognitive capacity is perceived as being a unique characteristic of hominins, as a consequence of the evolution of a large brain. However, in order to understand the evolution of modern human cognition and the cognitive capacities of our ancestors, one must distinguish between artefacts which reflect complex cognition, and those which reflect a more primitive cognition. Achieving this will allow an insight into the complexity of early hominins’ cognition, through a better understanding of the cognitive processes required to produce certain artefacts. Recently, observations of the behaviours of capuchins has called into question the attribution of two/three-million-year-old stone flakes to hominins (Proffitt et al. 2016, 1). Through stone-on-stone percussion, capuchins were able to produce stone flakes remarkably similar to those from the archaeological record (Figure 2), which are considered to be the earliest evidence of hominins producing tools (Proffitt et al. 2016, 2). These observations of primate behaviour have provided further support for the paradigm shift away from presuming the genus Homo is unique in creating sharp-edged flakes and tools (Proffitt et al. 2016, 3). Certainly, the earliest tool culture (Oldowan) demonstrates spatial coordination and planning, suggesting the Oldowan culture had evolved from earlier tool use (Haslam et al. 2009, 339). Further evidence for early tool use is derived from the observations of modern primates using a diverse range of tools habitually (McGrew 2010, 579). McGrew (2010) describes how chimpanzees are capable of using tool ‘sets’, such as five objects used in sequence to obtain honey. In addition, he outlines how chimpanzees use tool composites, such as a stone and anvil, and compound tools (several components combined into one working unit), such as inserting a wedge stone under an anvil. These types of tools were thought to be produced only by Homo, due to their relative complexity. The observations of primates producing tools may indicate that the common ancestor of modern humans and primates used tools (Haslam et al. 2009, 339).

Analogies with modern primates can also enable researchers to understand the process by which a characteristic evolved in modern humans. Although research has shown the gene mutation required for language (FOXP2) stabilised only 200,000 years ago (Enard et al. 2002, 871), this length of time is insufficient to explain the evolution of the complex cognitive apparatus which allows for spoken language (Zuberbühler 2003, 266). Modern primates have been shown to understand spoken English, use gestures to communicate (thus attaching meaning to gestures) (Savage-Rumbaugh et al.1986, 211; Matuzawa 2010, 5), and distinguish between different languages (Ramus et al. 2000, 351). This suggests that cognitive precursors to language were present in the last common ancestor (Zuberbühler 2003, 298), and thus resolves the inconsistency with the insufficient length of time for the evolution of language. Therefore, it is likely early hominins were able to use some form of language before the ability for spoken language was acquired (Zuberbühler 2003, 299).
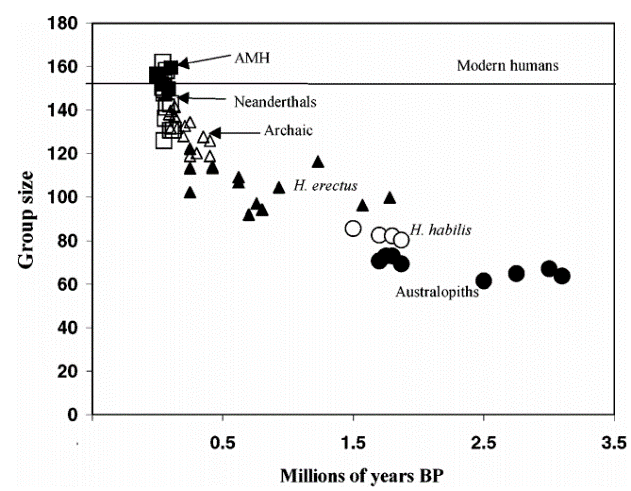
In addition, observations of modern primate social behaviour have given rise to the social brain theory (Dunbar 2003), which argues the evolution of abnormally large brains in primates and hominins was driven by increased social interactions. Dunbar (2003, 171) has shown the level of intentionality among modern primates and modern humans, with regard to theory of mind, correlates linearly with absolute frontal lobe volume. This implies that as brain size increased in hominins, the ability to socialise successfully with more individuals also increased (Figure 3). Therefore, through the comparison of modern primate and modern human behaviours, an insight into the maximum group sizes of hominins was achieved. Although this is not strictly an example of the use of primate analogies, it demonstrates how modern primate behaviour can enrich our understanding of early humans.
Social Structure
Evidence for early hominin social structure is elusive, therefore the use of primate analogies can provide an insight into the structure of these societies and how they evolved. Chimpanzees demonstrate intricate social behaviours, such as developing and reinforcing relationships (grooming with both non- and related individuals), hierarchical systems and an understanding of social (or cultural) ‘norms’ (Whiten et al. 1999, 685; Mitani et al. 2002, 10; Malone et al. 2012, 1264). These social behaviours appear to be integral to day-to-day life, for example grooming ensures social cohesion amongst primate groups (Aiello and Dunbar 1993, 185). Through observations of social grooming among modern primates, Aiello and Dunbar (1993, 185) suggest that early humans would have had to devote around 30 – 45% of their time to social grooming, in order to maintain large social group sizes (proposed in Figure 3). This significant proportion of time highlights the importance of social grooming to hominins. In addition, the capacity of modern primates to engage in these complex social interactions suggests the last common ancestor of hominins and great apes lived in groups consisting of numerous individuals (Malone et al. 2012, 1251).
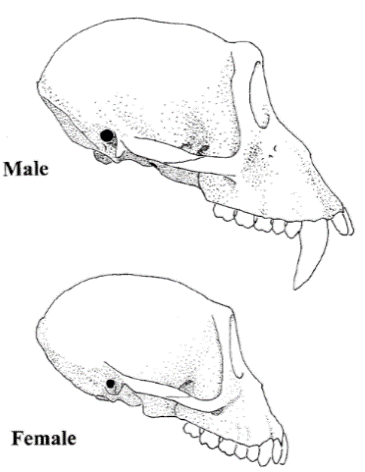
Furthermore, the morphological adaptations to social life amongst modern primates can be used to make inferences about aspects of hominin morphology. Modern primates, for example, have sexually dimorphic canines (Figure 4) which are used in displays of aggression and dominance, usually toward predators (Leutenegger and Kelly 1977, 126). Human evolution demonstrates a reduction in these canines; one can make inferences about the role of sexually dimorphic canines in early hominins and the factors which resulted in this reduction, through observing the role of these canines in modern primates. In addition, sexual dimorphism in body mass can also allow an insight into hominin lives. In modern primates, sexual dimorphism reflects social relationships; gorillas and orangutans have extreme sexual dimorphism, as a result of intense male-male competition to have control of females (Foley and Lee 1989, 904; Larsen 2003, 9103), whereas gibbons have little sexual dimorphism as the males are not in competition for mates. This can be used to understand sexual dimorphism (or lack of) in early humans. Sexual dimorphism in Australopithecus afarensis has been demonstrated to be similar to modern humans, suggesting there was little competition between males and that early hominin social behaviour may have been more like modern humans than previously thought (Larsen 2003, 9104; Reno et al. 2003, 9404).
Although modern primate behaviours are useful when it comes to understanding the diversity and intricacy of social behaviours among hominoids, one must tentatively use these comparisons. Interpretations of hominin social behaviour have previously been heavily reliant on chimpanzee models, and have thus led to assumptions involving closed male-dominant social networks with a lack of female alliances. However, as demonstrated by Parish and de Waal (2006) these assumptions are inappropriate and dismiss observations of another closely related primate, the bonobo. Amongst bonobos, females claim and control feeding, are violent toward males, and hunt and possess meat (Parish and de Waal 2006, 100). These activities in hominin behavioural models are usually attributed to males, without considering the possibility that females were the dominant sex. Thus, perceptions of hominin behaviour are inherently biased, which inevitably has had a negative impact on the understanding of human ancestors.
Cultural Behaviour
The cognitive ability to display culture is considered to be a decisively human characteristic, and debates exist regarding whether recent hominins, such as Neanderthals, were capable of cultural behaviours (Hayden 2012, 1). As defined by Freeman (2002), culture is:
“The total configuration of patterned activities performed by a society, including the materials used in or produced by those activities and the social units responsible for activity performance.”
Using this definition, certain behaviours indicate many modern primates are cognitively capable of culture. Culture allows for a group of individuals to work cohesively, and distinguish themselves from other such groups. Therefore, it requires advanced cognition to be able to create a set of behaviours which are shared with other individuals and considered unique to the group. Studies by Whiten et al. (1999) have observed variations in cultural behaviours in chimpanzees across Africa. Whiten et al. (1999, 685) argues that the variation in cultural behaviours observed in chimpanzees resembles that of human societies; the ability to produce culture is, therefore, no longer restricted to modern humans, opening up the possibility that early hominin ancestors exhibited cultural behaviours (Whiten et al. 2011, 939).
In addition, studies conducted by Lycett et al. (2009) have determined the prevailing method of cultural transmission among chimpanzee in order to understand how early hominins may have transmitted information. Similarities and differences in behaviour among chimpanzee groups studied appear to be consistent with the divergence of western and eastern subspecies, indicating that vertical cultural transmission (from one generation to the next) is the dominant process (Lycett et al. 2009, 344). Further, they demonstrated that females are more likely to pass on their genes if they adopt the cultural behaviours of a new group, restricting the success of horizontal transmission and suggesting that an affinity to conformist behaviour is not unique to modern humans (Whiten et al. 2005, 739; Lycett et al. 2009, 345).
Compassion and Morality
Through observing the moral behaviour of modern primates, one can understand how moral behaviour evolved in modern humans and the driving forces behind the emergence of this behaviour (Brosnan 2011, 23). Modern primates appear to display compassionate and moral behaviours to an extent; however they will also display behaviours which would be considered ‘selfish’ in a modern human context.
It has been demonstrated that primates have behavioural indicators for empathy. Behavioural copying (such as yawning when another yawns), is prevalent in modern humans and is considered vital to modern human’s ability to empathise. Higher-order primates also exhibit behavioural copying, which has been used to argue that they have the capacity to empathise with others, to some extent (De Waal 2008, 288). De Waal (2008, 284) uses the example of chimpanzees who upon hearing distress of another individual, will often feel distressed themselves (emotional contagion) and may even make efforts to comfort the individual in distress. The numerous parallels with primate empathetic behaviour with that of modern humans supports the notion that early hominins must have also been capable of these behaviours. In addition, Brosnan (2011) outlines how in certain experimental settings, primates appear to have a concept of inequity and fairness, for example a primate may refuse a reward for a task if their social partner does not receive an equal reward for the same task. This was used to suggest how the concept of inequity evolved in hominins; inequity has to be recognised, have a negative response in an individual, and be acted upon to rectify the inequity (Brosnan 2011, 28).
Adoption of orphaned juveniles amongst chimpanzees is another behaviour which suggests primates are capable of compassionate and moral behaviour (Spikins 2015, 86). Boesch et al. (2010) found that despite being a costly endeavour, with no apparent benefit to the adoptive parent nor any additional motivation behind doing so, chimpanzees would adopt both related and unrelated juveniles. This led to the conclusion that welfare for others is integral to chimpanzee groups (particularly the ones observed in the study, who had an increased risk of predators), and the tentative suggestion that the evolution of altruistic behaviour may have been a result of increased socio-ecological pressures.
To suggest primates are capable of altruistic and moral behaviours implies they are capable of a theory of mind not too dissimilar to that of modern humans. This has been a topic of debate since Premack and Woodruff’s (1978) paper, which speculated about a theory of mind in chimpanzees. It has been recently argued that primates are capable of a theory of mind (Krupenye et al. 2016), which may open up the possibility that early hominins (and perhaps the last common ancestor) were capable of this advanced level of cognition. With consideration of the numerous debates surrounding this topic, the implications of altruistic behaviours for a theory of mind in primates, and consequently the implications for the same capabilities in early hominins should be considered.
There are issues that arise with attributing primate behaviour as ‘altruistic’ or ‘moral’. One cannot understand the level of intentionality behind a primates’ actions, and so labelling actions in the same manner as human actions causes issues with distinguishing between the outcome of the action and the motivation. Primate A’s actions may have had positive repercussions for primate B, but this does not necessarily mean primate A intended to help primate B. Assuming that an intention to benefit other individuals exists in primates, and using this to further suggest hominins were capable of this level of altruism bases the behaviour of past hominins on too many assumptions and inherently causes issues in understanding early human behaviour. There is compelling evidence to suggest that perhaps higher-order primates can exhibit a level of altruism (which may have been similar to hominins), but one must use this evidence with caution particularly when using this as an analogy with early humans.
Conclusion
Primate analogies are inevitably useful to gain an insight into hominin behaviours, particularly when the material evidence is lacking. With regard to understanding the cognitive capacities of early humans, primate analogies are particularly beneficial to work by some researchers such as Dunbar (2003), Zuberbühler (2003) and Proffitt et al. (2016), and similarly can enrich the understanding of past social and cultural behaviours. However, these analogies must be used tentatively. Although useful, dependency on primate analogies with little additional evidence inevitably causes issues, and may lead to assumptions being drawn regarding early human behaviour which are inappropriate. Furthermore, in the past the use of primate analogies has caused issues with suggestions that hominin ancestors exhibited the ‘brutish’ and ‘primitive’ behaviour of apes (a particular prominent example of this is the depiction of Neanderthals in the early 20th century). Although these depictions of early humans may seem trivial, they are important in highlighting the extent one should apply primate analogies to the past, in order to prevent skewed interpretations of early humans. Therefore, it must be stressed that primate analogies should only provide an insight into early human behaviour, and it is unrealistic to assume that the observations of primate behaviour can be absolutely paralleled with hominin behaviour.
Bibliography
- Aiello, L.C. and Dunbar, R.I.M. (1993) ‘Neocortex Size, Group Size, and the Evolution of Language.’ Current Anthropology 34 (2): 184 – 193.
- Boesch, C., Bolé, C., Eckhardt, N. and Boesch, H. (2010) ‘Altruism in Forest Chimpanzees: The Case of Adoption.’ PLOS One 5 (1): 1 – 6.
- Brosnan, S. F. (2011) ‘An evolutionary perspective on morality.’ Journal of Economic Behaviour and Organisation 77: 23 – 30.
- De Waal, F.B.M. (2008) ‘Putting the Altruism Back into Altruism: The Evolution of Empathy.’ Annual Review of Psychology 59: 279 – 300.
- Dunbar, R.I.M. (2003) ‘The Social Brain: Mind, Language and Society in Evolutionary Perspective.’ Annual Review of Anthropology 32: 163 – 181.
- Enard, W., Przeworski, M. Fisher, S.E., Lai, C.S.L., Wiebe, V., Kitano, T., Monaco, A.P. and Pӓӓbo, S. (2002) ‘Molecular evolution of FOXP2, a gene involved in speech and language.’ Nature 418: 869 – 872.
- Foley, R.A. and Lee, P.C. (1989) ‘Finite social space, evolutionary pathways, and reconstructing hominid behavior.’ Science 243 (4893): 901 – 906.
- Freeman, L.G. (1978) The Analysis of Some Occupation Floor Distributions from Earlier and Middle Palaeolithic Sites in Spain. In: Freeman, L.G. (ed.) Views of the Past: Essays in Old World Prehistory and Paleanthropology. De Gruyter Mouton.
- Haslam, M., Hernandez-Aguilar, A., Ling, V., Carvalho, S., de la Torre, I., DeStefano, A., Du, A., Hardy, B., Harris, J., Marchant, L., Matsuzawa, T., McGrew, W., Mercader, J., Mora, R., Petraglia, M., Roche, H., Visalberghi, E. and Warren, R. (2009) ‘Primate Archaeology.’ Nature 460: 339 – 344.
- Krupenye, C., Kano, F., Hirata, S., Call, J. and Tomasello, M. (2016) ‘Great apes anticipate that other individuals will act according to false beliefs.’ Science 354 (6308): 110 – 114.
- Larsen, C.S. (2003) ‘Equality for the sexes in human evolution? Early hominid sexual dimorphism and implications for mating systems and social behavior.’ Proceedings for the National Academy of Science 100 (16): 9103 – 9104.
- Leutenegger, W. and Kelly, J.T. (1977) ‘Relationship of Sexual Dimorphism in Canine Size and Body Size to Social, Behavioral and Ecological Correlates in Anthropoid Primates.’ Primates 18 (1): 117 – 136.
- Lycett, S.J., Collard, M. and McGrew, W.C. (2009) ‘Cladistic analyses of behavioural variation in wild Pan troglodytes: exploring the chimpanzee culture hypothesis.’ Journal of Human Evolution 57: 337 – 349.
- Malone, N., Fuentes, A. and White, F.J. (2012) ‘Variation in the Social Systems of Extant Hominoids: Comparative Insight into the Social Behavior of Early Hominins.’ International Journal of Primatology 33 (6): 1251 – 1277.
- Matuzawa, T (2010) The Chimpanzee Mind: Bridging Fieldwork and Laboratory Work. In: Lonsdorf, E., Ross, S.R. and Matuzawa, T. (eds.) The Mind of the Chimpanzee: Ecological and Experimental Perspectives. University of Chicago Press.
- McGrew, W.C. (2010) ‘Chimpanzee technology.’ Science 328 (5978): 579 – 580.
- Mitani, J.C., Watts, D.P. and Muller, M.N. (2002) ‘Recent Developments in the Study of Wild Chimpanzee Behavior.’ Evolutionary Anthropology 11 (1): 9 – 25.
- Parish, A.R. and de Waal, F.B.M. (2006) ‘The Other “Closest Living Relative.” How Bonobos (Pan paniscus) Challenge Traditional Assumptions about Females, Dominance, Intra- and Intersexual Interactions, and Hominid Evolution.’ Annals of New York Academy of Sciences 907 (1): 97 – 113.
- Plavcan, J.M. (2001) ‘Sexual Dimorphism in Primate Evolution.’ Yearbook of Physical Anthropology 44: 25 – 53.
- Premack, D. and Woodruff, G. (1978) ‘Does the chimpanzee have a theory of mind?’ The Behavioural and Brain Sciences 4: 515 – 526.
- Proffitt, T., Luncz, L.V., Falótico, T., Ottoni, E.B., de la Torre, I. and Haslam, M. (2016) ‘Wild monkeys flake stone tools.’ Nature 539: 85 - 88.
- Ramus, F., Hauser, M.D., Miller, C., Morris, D. and Mehler, J. (2000) ‘Language Discrimination by Human Newborns and by Cotton-Top Tamarin Monkeys.’ Science 288 (5464): 349 – 351.
- Reno, P.L., Meindl, R.S., McCollum, M.A. and Lovejoy, C.O. (2003) ‘Sexual dimorphism in Australopithecus afarensis was similar to that of modern humans.’ Proceedings for the National Academy of Science 100 (16): 9404 – 9409.
- Savage-Rumbaugh, S., McDonald, K., Sevcik, R.A., Hopkins, W.D., and Rupert, E. (1986) ‘Spontaneous symbol acquisition and communicative use by pygmy chimpanzees (Pan paniscus).’ Journal of Experimental Psychology 115 (3): 211 – 235.
- Spikins, P. (2015) How Compassion Made Us Human: The Evolutionary Origins of Tenderness, Trust and Morality. Pen and Sword.
- Whiten, A., Goodall, J., McGrew, W.C., Nishida, T., Reynolds, V., Sugiyama, Y., Tutin, C.E.G.,
- Wrangham, R.W. and Boesch, C. (1999) ‘Cultures in Chimpanzees.’ Nature 399: 682 – 685.
- Whiten, A., Hinde, R. A., Laland, K. N., and Stringer, C.B. (2011) ‘Culture evolves.’ Philosophical Transactions of the Royal Society of London B 366 (1567): 938 – 948.
- Whiten, A., Horner, V. and de Waal, F.B.M. (2005) ‘Conformity to cultural norms of tool use in chimpanzees.’ Nature 437: 737 – 740.
- Zuberbühler, K. (2003) ‘Referential Signaling in Non-Human Primates: Cognitive Precursors and Limitations for the Evolution of Language’ Advances in the Study of Behavior 33: 265 – 307.